XXI.4.2.2.1 The most important prezygotic reproductive isolation mechanisms involve spatial, temporal, ethological and morphological isolation
Spatial isolationof a sympatric species is an important prezygotic reproductive isolation mechanism (RIM). If two species inhabit different biotopes in a common range or utilize different plants for food within a single biotope, their members will encounter one another (and thus reproduce) far less frequently than members of the same species. Temporal isolation functions similarly. If two species occur in the same territory but are active at different times of the day, they need never meet. Temporal and spatial isolation can be included amongst both external and internal prezygotic reproductive barriers, as they can be substantially affected both by external factors and by genetically determined differences in the behavior of individuals in the two differentiating species.
Internal prezygotic RIM include ethological isolation mechanisms, which encompass specific patterns of behavior, through which the members of a single species communicate prior to reproduction or during its progress. Even very close species can differ substantially in the character and timing of these patterns of behavior, where differences in behavior can very effectively prevent interspecific crossing. Mutual seeking out of the members of the opposite sex for the purpose of reproduction is very frequently accompanied by unilateral or bilateral exchange of acoustic, chemical and/or mechanical signals. The receptors of these signals are generally very specific. For example, the receptors of acoustic signals do not register sounds whose frequency differs only slightly from that of the species-specific signal. Ethological isolation mechanisms can also be important for flowering plants. In this case, pollinators also participate in these mechanisms. The flower sends out species-specific optical or chemical signals, attracting certain species of pollinators. If two close species of plants differ in their signals, they can also differ in their spectra of pollinators. This can substantially reduce pollination of the oocytes by the pollen of a foreign species.
Morphological isolationis another type of prezygotic isolation mechanism. Put simply, the male and female sex organs of two individuals need not fit together (Sota & Kubota 1998). Great importance was attributed to morphological isolation in the past, especially amongst arthropods. It is known that even very closely related species of arthropods have very different shapes of their copulation organs. In a great many cases, related species can be differentiated only on the basis of the morphology of these organs. Frequently various complicated protrusions are formed on the copulation organs, so that they naturally evoke the idea of a sort of lock and key capable of ensuring an effective interspecific reproductive isolation barrier. However, at the present time, it seems that the evolutionary reasons for the formation and rapid development of complicated copulation organs in insects will lie elsewhere. Mechanical reproduction barriers are mostly rather ineffective and, in addition, the copulation organs also rapidly diverge in allopatric species {11021}. It is highly probable that this could entail a co-evolutionary battle between males and females over control of the fate of the ejaculate inside the female body. The female frequently has quite different biological interests in how to manage the sperm than those of the male that mated with her. While the female attempts within the context of cryptic female choice to select, from the sperm obtained from various males, that from the best-quality male, or attempts to ensure that the individual oocytes are fertilized by the sperm of different males, it is, to the contrary, advantageous for the male if the female employs his sperm to fertilize the greatest number of oocytes, regardless of his own genetic quality or the genetic quality of the female. In this co-evolutionary battle, the two sexes employ various ethological, chemical and mechanical weapons and counter-weapons, through which both sexes attempt to achieve their contradictory goals. Because of the co-evolutionary character of this battle, the evolution of the individual mechanisms and thus of the corresponding morphological structures is extremely fast. If, in the experiment, we prevent the males or females from responding to the evolutionary moves of the members of the opposite sex, they lose the co-evolutionary battle in a few generations and the results of copulation begin to be unilaterally advantageous for the members of the other sex (Fig. XXI.6).
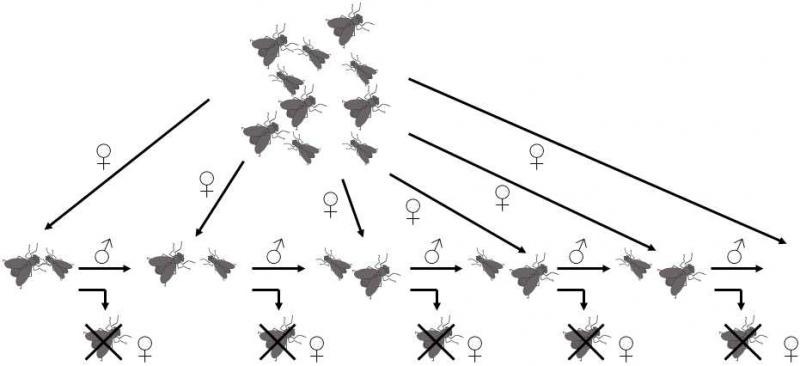
Fig. XXI.6 Coevolutionary battle between males and females. Scheme of an experiment in which females were prevented from responding evolutionarily to the evolutionary moves of males. In each generation, the females in the experimental vessel were replaced by “naive” females from the stock breeding. After 30 generations, the males had 25% greater fitness compared with the control males, while the females exhibited significantly greater mortality after copulating with experimental males than after copulating with control males.
For example, in experiments, the males of drosophila were kept for long periods of time in the cultivation vessel, while the females were removed in each generation and replaced by “naive” females (i.e. females obtained from a different breeding). Within 30 generations, the experimental males already had 24% greater fitness compared with the control males. This was caused primarily by the fact that they were capable of copulating more frequently with females that formerly copulated with a different male and also by the fact that the females that copulated with them did not subsequently copulate with a different male. The means, or rather the chemical instruments, through which the males achieved this, simultaneously damaged the females in some way. Females that copulated with the experimental males exhibited substantially higher mortality than females that copulated with control males (Rice 1996). It is highly probable that the present-day complicated morphology of copulation organs is a result of just such a co-evolutionary battle and its interspecific differences are only a side effect of the accelerated evolution of these structures.
Gamete incompatibility represents another barrier preventing interspecific crossing. This barrier is especially important in species whose members release their gametes into the open environment, where they actively seek out one another. In many cases, the gametes seek one another through species-specific chemical attractants. It very often happens that the microgametes require specific molecules that enable them to penetrate into the macrogametes of their own species. It is significant that the surface proteins of a gamete are amongst the molecules with the greatest rate of evolutionary accumulation of nonsynonymous changes. Gamete incompatibility is very important in plants. The pollen of foreign species very frequently reaches the stigma of flowers. The pollen mostly germinates, but the growth of the pollen tubes is mostly slow and the tubes mostly do not reach the oocytes.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.