XXI.4.3.6 Haldane’s rule is sometimes explained by the action of ultraselfish genetic elements that accumulate primarily in the nonrecombining parts of the Y and W allosomes
The hypothesis of revolt of ultraselfish genes is based on the idea that, in the area of the genome in which recombination does not occur, ultraselfish genetic elements can accumulate and spread in the population by one of the mechanisms of evolutionary drive (see Chap. VI). Generally, the cooperation of several genes is required for their spreading, say gene A, whose product would damage the chromosomes derived from the other parent, and gene B, whose products would protect the chromosomes from the same gene set and thus from the same parent, against the action of the product of gene A (Fig. XXI.11). If both genes are located on an autosome, they can be separated in the progeny as a result of genetic recombination, understandably with catastrophic results for spreading of gene A – in the next generation, it will damage the chromosomes of both chromosome sets and thus basically commit genetic suicide. In contrast, the genes in the nonrecombining DNA sections, i.e. particularly in unpaired sex chromosomes (allosomes) Y and W, are always transferred together and thus form a sort of supergene. They can form coalitions and spread in the gene pool of the population even at the expense of the average viability of its members.
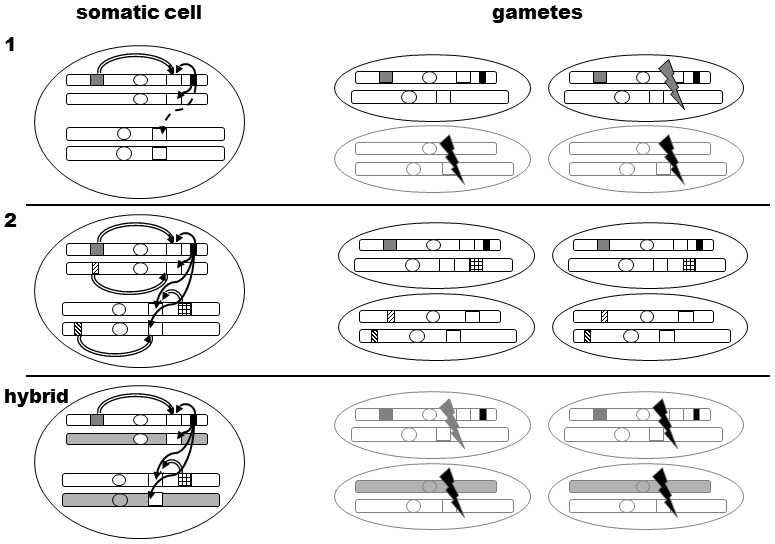
Fig. XXI.11 Mechanism of the Haldane rule according to the hypothesis of the revolt of ultraselfish genetic elements. In the first phase, a chromosome containing pairs of genes that are in genetic linkage appears and is fixed in time in the species. This is an ultraselfish gene (white rectangle) producing a toxin that destroys primarily the homologous chromosome containing a different allele at the particular site (black arrow), and also a gene (grey rectangle) that renders the chromosome containing the ultraselfish gene resistant to the action of this toxin (white arrow) and protects it against the toxin. Because of the requirement on the existence of genetic linkage between the two genes, an Y-chromosome at which recombinations do not occur is the best candidate for the formation of ultraselfish genes and the chief “target for attack” is the X-chromosome. The toxin damages autosomes with somewhat lesser effectiveness (dashed arrows). The action of the toxin results in the individual having reduced fertility; however, all its gametes contain the ultraselfish gene, to be more exact the ultraselfish allele, at the particular locus. In the second phase, various gene neutralizers are fixed at all the other chromosomes (checked and hatched rectangles), making these chromosomes resistant to the action of the toxin, or alleles, that lost their target sites on which the toxin acted, are fixed here. The fertility of carriers of an ultraselfish gene is then fully renewed and the presence of this gene is no longer manifested. However, if interspecies hybridization occurs, the ultraselfish gene finds itself in the presence of nonresistant chromosomes and begins to be manifested (killing or damaging gametes is depicted as black or grey “thunderbolts”). If the given species were to have only two sex chromosomes and one pair of autosomes in the nucleus, as depicted for simplicity in the figure, a quarter of the sperm would be functional. However, in actual fact, there are far more pairs of autosomes. It is sufficient for dysfunctionality of the gamete if the gene set of the gamete contains at least one sensitive autosome derived from a foreign species. As several ultraselfish elements could be gradually fixed on the Y-chromosomes of the same species, their joint action on the autosomes could be manifested in sterility of hybrids of the heterogametic sex.
The parliament of genes model (Leigh 1972)assumes that the spreading of such ultraselfish genes in the population is very rapidly prevented by the spreading of some other gene, to be precise some alleles of some other gene, which are capable of neutralizing the function of the ultraselfish gene. Genes on chromosome Y can be readily inactivated, for example by integration of a transposon or retrotransposon. This could be connected with observed accumulation of transposons, in humans primarily retrotransposons, in nonrecombining Y-chromosome areas (Erlandsson, Wilson, & Paabo 2000). Within a species, ultraselfish genes that are located on allosomes are not greatly manifested as they are “held in check” by the appropriate neutralizer genes, located on the other chromosomes. However, as soon as an allosome in a hybrid finds itself in the presence of a foreign gene set, the ultraselfish genes can begin to act and damage both the fertility and viability of their bearers (Tao, Hartl, & Laurie 2001). The presence of its own chromosome set with the relevant neutralizer genes need not necessarily protect a hybrid against the action of an ultraselfish gene, as neutralizer genes can, for example, be capable of protecting only their own chromosome. For example, these can be alleles that have lost the target site for the product of the ultraselfish gene. Another possibility is that the neutralizer can act (for example by protecting the target sites on the chromosome of their own chromosome set by methylation) during the progress of gametogenesis, i.e. sooner than the chromosomes in the zygote come into interaction with the products of the ultraselfish gene.s
The results of some experiments and a number of observations in nature support the action of ultraselfish genes in the formation of postzygotic interspecific barriers (Orr & Presgraves 2000). It has been found primarily in flora that one of the potential consequences of interspecific hybridization and also, e.g., polyploidization (Wendel 2000)consists in the activation of genetic elements of the transposon type, their cutting out and insertion into new sites (Comai 2000). Compared to animals, plants have a far more dynamic genome and are frequently capable of repairing damage to their DNA occurring as a consequence of increased transposon activity. Reparation is frequently accompanied by fundamental restructuring of the genome and this restructuring can also be substantially manifested in the phenotype of hybrid flora and their progeny. It is probable that the passivity of transposons in normal plants is a result of neutralizer genes that are gradually fixed in the gene pool of the plant as a result of transposone selection pressure. Neutralizer genes need not occur in the genome of a foreign species or their functioning in a genome containing the chromosomes of a foreign species are greatly limited so that they are not capable of completely controlling the transposons activity.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.